In diesem Abschnitt werden zunächst die durchgeführten Messungen zusammengestellt. Danach werden die optimalen Parameter der elektrischen Stimulation bestimmt. Am Ende werden die einzelnen Messungen und gewonnene Daten ausführlich vorgestellt.
Mit der Organbadanlage ohne Schrittmotorerweiterung lässt sich nur
die isometrische Messung durchführen. Bei dieser Messung wird die
Länge der Probe über die Mikrometerschraube voreingestellt und
dann während einer Messung nicht mehr verändert (isometrisch). Bei
Aktivierung des Muskels durch elektrische Stimulation wird eine
Kontraktionskraft ![]() am Kraftaufnehmer gemessen (siehe
Abbildung 19). Die maximale Kontraktionskraft
am Kraftaufnehmer gemessen (siehe
Abbildung 19). Die maximale Kontraktionskraft
![]() , die der Muskel nach Aktivierung zusätzlich zur
Ruhespannung
, die der Muskel nach Aktivierung zusätzlich zur
Ruhespannung ![]() noch erzeugen kann, ist von der
voreingestellten Längung
noch erzeugen kann, ist von der
voreingestellten Längung ![]() abhängig (siehe Abbildung
20).
abhängig (siehe Abbildung
20).
Bei der passiven Messung wird der Muskel sprunghaft verlängert
oder verkürzt, d.h. die Längenänderungen von ![]() erfolgen in
einem Zeitraum von weniger als
erfolgen in
einem Zeitraum von weniger als ![]() . Der sich daraufhin
einstellende Ausgleichsprozess
. Der sich daraufhin
einstellende Ausgleichsprozess ![]() wird aufgezeichnet (siehe
Abbildung 16). Die sich am Ende dieses
Ausgleichsprozesses einstellende Kraft
wird aufgezeichnet (siehe
Abbildung 16). Die sich am Ende dieses
Ausgleichsprozesses einstellende Kraft ![]() ist von der
Länge des Muskels abhängig (siehe Abbildung 17).
ist von der
Länge des Muskels abhängig (siehe Abbildung 17).
Durch die Schrittmotoreinheit ist nun eine isotone Messung
möglich. Bei dieser werden die Kraft konstant gehalten (isoton)
und die bei einer Aktivierung sich einstellende Verkürzung des
Muskels aufgezeichnet (siehe Abbildung 23). Die
maximale Kontraktion
![]() , um die der Muskel noch
zusätzlich zur Ruhelänge
, um die der Muskel noch
zusätzlich zur Ruhelänge ![]() kontrahiert, hängt dabei von der
konstant zuhaltenden Kraft
kontrahiert, hängt dabei von der
konstant zuhaltenden Kraft ![]() ab (siehe Abbildung
25). Aus den Längen
ab (siehe Abbildung
25). Aus den Längen ![]() können die
Kontraktionsgeschwindigkeiten
können die
Kontraktionsgeschwindigkeiten ![]() berechnet werden. Die
maximale Kontraktionsgeschwindigkeit
berechnet werden. Die
maximale Kontraktionsgeschwindigkeit ![]() hängt dabei auch
von der Kraft ab (siehe Abbildung 26).
hängt dabei auch
von der Kraft ab (siehe Abbildung 26).
Bei der passiven Messung werden die Soll-Kraft der
Schrittmotorregelung innerhalb von ![]() sprunghaft verändert
und die sich daraufhin einstellende Längenänderung
sprunghaft verändert
und die sich daraufhin einstellende Längenänderung ![]() aufgezeichnet (siehe Abbildung 22). Die sich am Ende
dieses Ausgleichsprozesses einstellende Länge
aufgezeichnet (siehe Abbildung 22). Die sich am Ende
dieses Ausgleichsprozesses einstellende Länge ![]() ist von
der Kraft abhängig.
ist von
der Kraft abhängig.
Diese Zusammenhänge sind in Tabelle 1 dargestellt.
|
Zur Bestimmung des aktiven Verhaltens des Muskels ist es nötig,
den Muskel elektrisch so zu reizen, daß er möglichst stark
kontrahiert und dabei keinen Schaden nimmt. Am Signalgenerator
sind Dauer und Stromstärke der Signale einstellbar. Bei zu kleiner
Reizdauer oder Amplitude wird der Muskel nicht ausreichend
aktiviert; wo hingegen bei zu dauerhafter und starker Reizung der
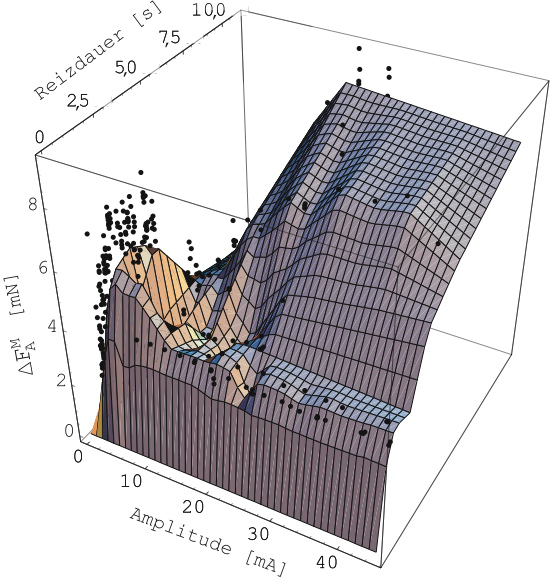
Muskel Schaden nimmt. In Abbildung 14 ist die
maximale Kraft ![]() bei isometrischer Kontraktion über
die Reizparameter Dauer und Amplitude aufgetragen (Probe 1-A).
Dabei ist durch über 500 Einzelmessungen (schwarze Punkte) eine
Ausgleichsfläche gezeichnet.
bei isometrischer Kontraktion über
die Reizparameter Dauer und Amplitude aufgetragen (Probe 1-A).
Dabei ist durch über 500 Einzelmessungen (schwarze Punkte) eine
Ausgleichsfläche gezeichnet.
Deutlich zu erkennen ist bei kleinen Amplituden (![]() ) und
kurzen Reizlängen (
) und
kurzen Reizlängen (![]() ) eine relativ starke Kontraktion. Nur
bei sehr viel größere Amplituden und längere Reizdauern
kontrahiert der Muskel noch stärker. Die nachfolgenden Messungen
des aktiven Verhaltens werden mit Reizen der Dauer von
) eine relativ starke Kontraktion. Nur
bei sehr viel größere Amplituden und längere Reizdauern
kontrahiert der Muskel noch stärker. Die nachfolgenden Messungen
des aktiven Verhaltens werden mit Reizen der Dauer von ![]() bis
bis
![]() und Amplituden von
und Amplituden von ![]() bis
bis ![]() durchgeführt.
durchgeführt.
Die Deutlichkeit, mit der das oben erwähnte relative Maximum bei kurzer Reizdauer und kleiner Reizamplitude hervortritt, ist erstaunlich. Die maximale Kontraktionskraft nach elektrischer Stimulation mit Rechteckimpulsen gleicher Amplitude und zunehmender Länge bzw. gleicher Länge und zunehmender Amplitude nimmt im allgemeinen in Form einer Sättigungskurve monoton zu. Es ist zu bedenken, dass der Muskel wahrscheinlich ein Gedächtnis für bereits erfolgte Stimulationen besitzt: wurde er z.B. zu stark gereizt, wird sich dies auf die folgenden Reaktionen auswirken. Aus der Sicht der Reiz-Geschichte, die der Muskel erfuhr, ist Abbildung 14 problematisch, liefert aber Anhaltspunkte für eine grobe Orientierung.
Es soll hier die passive Kraftentwicklung des Muskels bei Längung
untersucht werden. Die Abhängigkeit der Kraftentwicklung von der
Längung wird durch die Kraft-Längenkurve ![]() angegeben.
angegeben.
Abbildung 15 zeigt einen Ausschnitt aus den
Messdaten (Probe 2-B). Es wurde Kraft über Zeit aufgetragen. Die
Abtastung erfolgte mit ![]() . Lokale Kraftmaxima entsprechen
einer Längung der Probe und lokale Minima einer Verkürzung. Die
Längenänderung erfolgte manuell über eine Mikrometerschraube am
Organbad. Die Einstellungen erfolgten ruckartig und wurden
jedesmal notiert. Eine typische Längenänderung pro Schritt ist
. Lokale Kraftmaxima entsprechen
einer Längung der Probe und lokale Minima einer Verkürzung. Die
Längenänderung erfolgte manuell über eine Mikrometerschraube am
Organbad. Die Einstellungen erfolgten ruckartig und wurden
jedesmal notiert. Eine typische Längenänderung pro Schritt ist
![]() bei einer Gesamtlänge der Probe 2-B von
bei einer Gesamtlänge der Probe 2-B von ![]() .
.
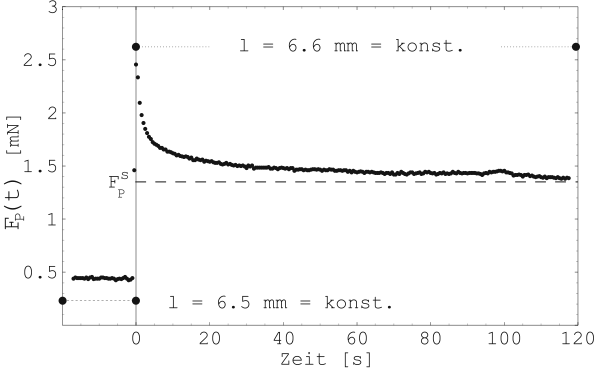
Abbildung 16 zeigt vergrößert einen einzelnen
Relaxationvorgang nach sprunghafter Längung der Gewebeprobe von
![]() auf
auf ![]() . Der Zeitnullpunkt fällt nun mit der
Längung der Probe zusammen.
. Der Zeitnullpunkt fällt nun mit der
Längung der Probe zusammen.
|
An Abbildung 16 sieht man, daß auch noch lange
Zeit nach der ruckartigen Längung die Probe nachgibt. Zwei Fragen
sind in diesem Zusammenhang von Interesse: Zum einen die Frage
nach dem zeitlichen Verhalten (![]() -Kurve), wie das Gewebe
nachgibt, und zum anderen nach dem stabilen Endzustand
(
-Kurve), wie das Gewebe
nachgibt, und zum anderen nach dem stabilen Endzustand
(![]() ) des Gewebes.
) des Gewebes.
Um aus den gemessenen ![]() -Kurven den stabilen Endzustand
-Kurven den stabilen Endzustand
![]() bestimmen zu können, wird eine in Abschnitt
7.2 beschriebene nichtlineare Regression durchgeführt.
Der Endzustand
bestimmen zu können, wird eine in Abschnitt
7.2 beschriebene nichtlineare Regression durchgeführt.
Der Endzustand ![]() entspricht dabei dem zu schätzenden
Parameter
entspricht dabei dem zu schätzenden
Parameter ![]() aus den Gleichungen 3 und 4
in Abschnitt 7.2.1.
aus den Gleichungen 3 und 4
in Abschnitt 7.2.1.
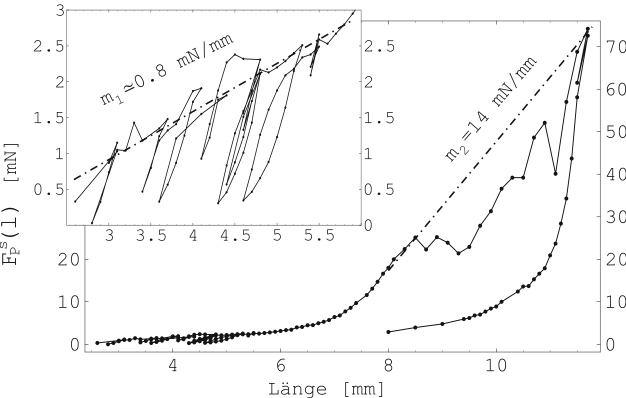
Abbildung 17 zeigt die so ermittelten Kräfte für ![]() in Abhängigkeit von der jeweiligen Längung der Probe.
Nacheinander gemessene Werte sind verbunden.
in Abhängigkeit von der jeweiligen Längung der Probe.
Nacheinander gemessene Werte sind verbunden.
|
In Abschnitt 2.4.2 wurde darauf hingewiesen, daß ein
überlineares Kraft-Längen-Verhalten für Muskelgewebe zu erwarten
ist. Das wird durch diese Messung bestätigt. Die in Abbildung
17 angedeuteten Ausgleichsgeraden dienen der
Abschätzung linear-elastischer Verhältnisse bei kleinen und
großen Längungen des Gewebes. Hier ergibt sich etwa der Faktor
20. Der übergangsbereich liegt etwa bei der ![]() -fachen
Ruhelänge.
-fachen
Ruhelänge.
Es soll hier die Kontraktionsfähigkeit des Gewebes bei
verschiedenen Längungen der Probe und gleichen
Stimulationsparametern untersucht werden. Die Abhängigkeit der
Kontraktionskraftentwicklung von der Längung wird durch die
isometrische Kraft-Längenkurve
![]() angegeben.
angegeben.
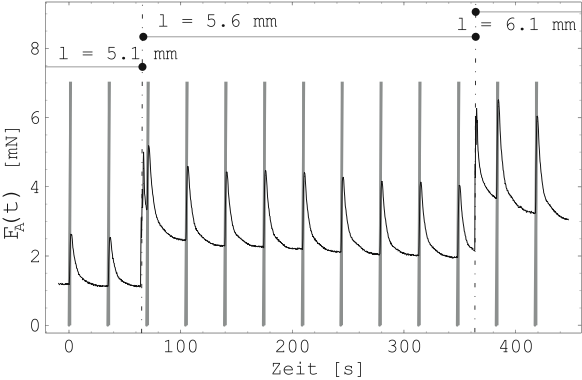
Abbildung 18 zeigt die Kontraktion des Gewebes
(schwarze Linie) bei elektrischer Stimulation (graue Rechtecke)
über der Zeit aufgetragen (Probe 3-C). Zum Zeitpunkt ![]() und
und
![]() wurde das Gewebe manuell nachgespannt. Die einzelnen
Längungen des Gewebes wurden jeweils notiert. Insgesamt wurden
etwa 300 Kontraktionen aufgezeichnet. Die elektrische Stimulation
ist für alle Kontraktionen ein Rechteckimpuls der Dauer 2 Sekunden
und mit der Amplitude von
wurde das Gewebe manuell nachgespannt. Die einzelnen
Längungen des Gewebes wurden jeweils notiert. Insgesamt wurden
etwa 300 Kontraktionen aufgezeichnet. Die elektrische Stimulation
ist für alle Kontraktionen ein Rechteckimpuls der Dauer 2 Sekunden
und mit der Amplitude von ![]() . Die Zeit zwischen den
einzelnen Rechteckimpulsen beträgt etwa 30 Sekunden. Während der
einzelnen Längungen wurde die Probe zwischen 10 und 20 mal
stimuliert.
. Die Zeit zwischen den
einzelnen Rechteckimpulsen beträgt etwa 30 Sekunden. Während der
einzelnen Längungen wurde die Probe zwischen 10 und 20 mal
stimuliert.
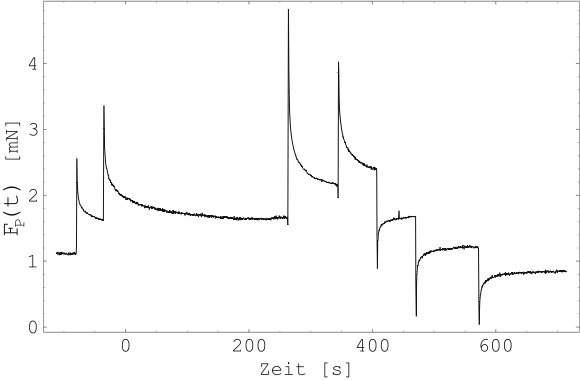
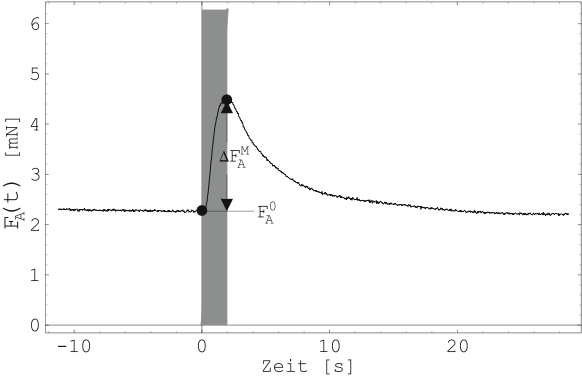
Abbildung 19 zeigt einen Einzelimpuls aus Abbildung
18. Die Ruhekraft ![]() des Muskels direkt
vor der Stimulation und die maximale zusätzliche Kraftentwicklung
des Muskels direkt
vor der Stimulation und die maximale zusätzliche Kraftentwicklung
![]() nach erfolgter Stimulation sind eingezeichnet.
Beide sind von der Länge des Muskels abhängig.
nach erfolgter Stimulation sind eingezeichnet.
Beide sind von der Länge des Muskels abhängig.
|
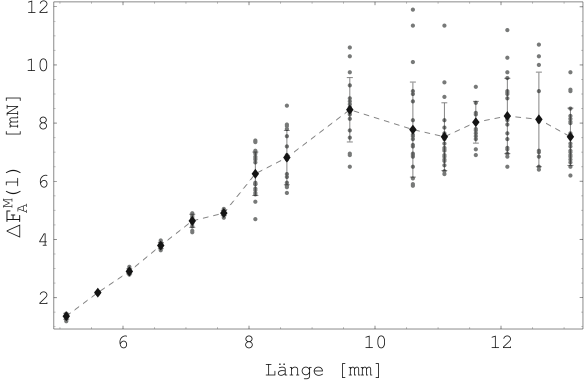
Die Abbildungen 20 zeigt die Mittelwerte und
Standardabweichungen der Kraftentwicklung
![]() der
Probe 3-C bei elektrischer Stimulation für verschiedene Längungen.
Die Abbildung 21 zeigt die Ruhekräfte
der
Probe 3-C bei elektrischer Stimulation für verschiedene Längungen.
Die Abbildung 21 zeigt die Ruhekräfte ![]() .
.
|
Die Abbildung 20 kann mit den Abbildungen 3 und 5 verglichen werden. Dabei ist auf die Skalierung der Längenachsen zu achten. Die Existenz eines Kraftmaximums bei bestimmter Muskellänge (hier zwei bis dreifache Ausgangslänge) wird nur schwach bestätigt.
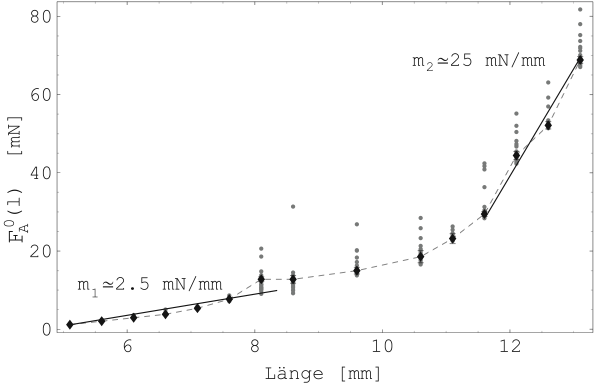
Die Abbildung 21 ist eigentlich auch eine Kraft-Längen-Kurve des passiven Muskels und damit mit Abbildung 17 vergleichbar. Wie in Abschnitt 6.3.3 ergeben sich die Verhältnisse der Elastizitätsmodule für kleine und große Längen zu etwa 1:10. Der übergangsbereich liegt auch bei etwa dem 2.5-fachen der Ausgangslänge des Muskels.
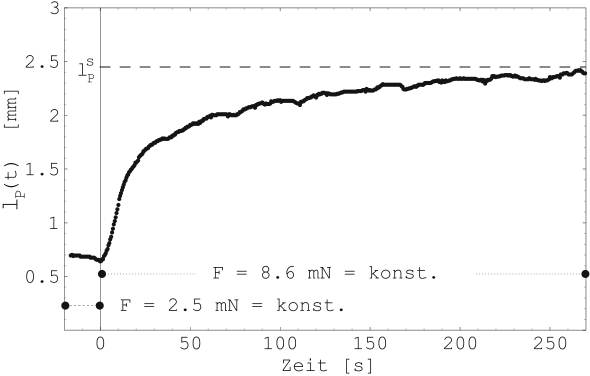
Bei dieser Messung werden die Kraft des Muskels sprunghaft
verändert und die sich daraufhin einstellende Längenveränderung
des Muskels ![]() aufgezeichnet (Kriechen). Abbildung
22 zeigt einen solchen Ausgleichsprozess (Probe 4-D).
aufgezeichnet (Kriechen). Abbildung
22 zeigt einen solchen Ausgleichsprozess (Probe 4-D).
Da diese Messungen gegenüber der isometrischen Messung des passiven Verhaltens keine neuen Informationen über die mechanischen Eigenschaften des Muskels liefert, erfolgen keine weiteren Messungen dieser Art.
Bei der isotonen Messung werden die Kraft konstant gehalten und
die bei einer Aktivierung notwendige Längenänderung des Muskels
![]() aufgezeichnet und deren zeitliche Ableitung
aufgezeichnet und deren zeitliche Ableitung ![]() bestimmt.
bestimmt.
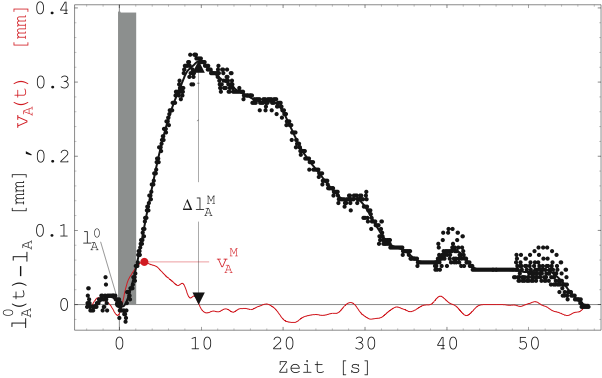
Abbildung 23 zeigt den zeitlichen Verlauf der vom
Schrittmotor nachgeregelten Probenlänge (schwarze Punkte, Probe
5-E). Rot eingezeichnet ist die rechnerisch ermittelte
Verkürzungsgeschwindigkeit mit Maximum ![]() . Damit die
numerische Differentiation brauchbare Werte liefert, wurden die
Orginaldaten geglättet (schwarze Linie).
. Damit die
numerische Differentiation brauchbare Werte liefert, wurden die
Orginaldaten geglättet (schwarze Linie).
|
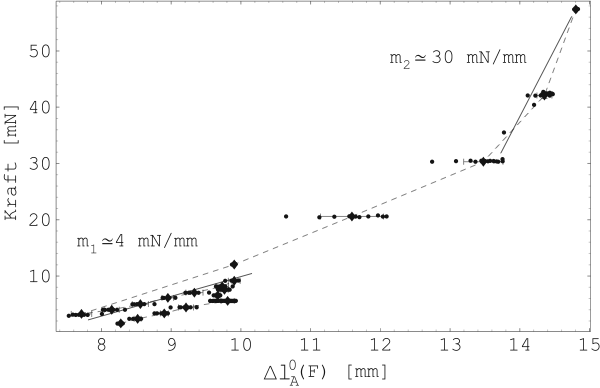
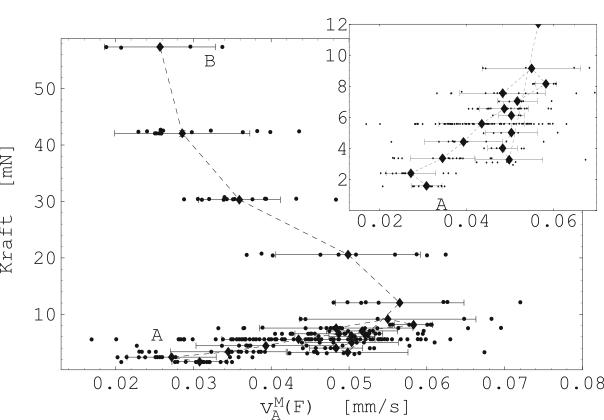
Abbildung 25 zeigt nun die ermittelten maximalen
Verkürzungen
![]() mit den dazugehörigen konstant
gehaltenen Kräften (Probe 5-E). Es wurden zwischen 10 und 40
Kontraktionen für jede eingestellte Vorspannung nacheinander
durchgeführt. Die Mittelwerte dieser Kontraktionen sind in der
Messreihenfolge durch eine gestrichelte Linie verbunden. A
markiert den Anfang und B das Ende (siehe Abbildung
25). Abbildung 24 zeigt die
dazugehörigen Ruhelängen
mit den dazugehörigen konstant
gehaltenen Kräften (Probe 5-E). Es wurden zwischen 10 und 40
Kontraktionen für jede eingestellte Vorspannung nacheinander
durchgeführt. Die Mittelwerte dieser Kontraktionen sind in der
Messreihenfolge durch eine gestrichelte Linie verbunden. A
markiert den Anfang und B das Ende (siehe Abbildung
25). Abbildung 24 zeigt die
dazugehörigen Ruhelängen ![]() .
.
|
|
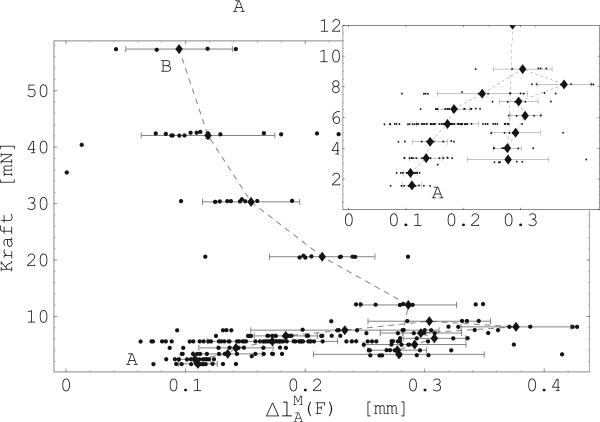
Abbildung 26 zeigt nun die ermittelten, maximalen Verkürzungsgeschwindigkeiten mit den dazugehörigen, konstant gehaltenen Kräften (Probe 5-E). Es wurden zwischen 10 und 40 Kontraktionen für jede eingestellte Vorspannung nacheinander durchgeführt. Die Mittelwerte dieser Kontraktionen sind in der Messreihenfolge durch eine gestrichelte Linie Verbunden (siehe Abbildung 26). A markiert den Anfang und B das Ende .
|
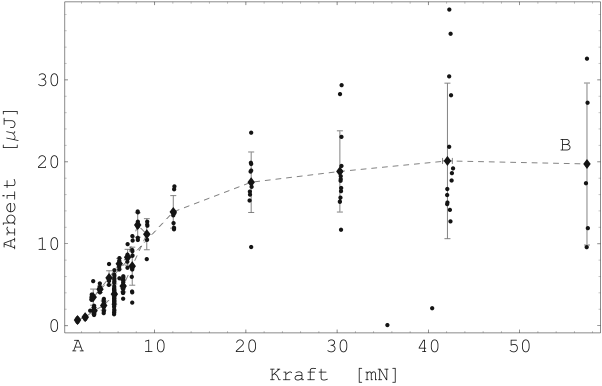
Bei der isometrischen Messung leistet der Muskel keine äußere mechanische Arbeit. Bei der isotonischen Messung lässt sich die geleistete, mechanische Arbeit jedoch aus dem Produkt der konstant gehaltenen Kraft und der nachgeregelten Länge berechnen. Die Abbildung 27 zeigt die geleistete, mechanische Arbeit in Abhängigkeit der Kraft (Probe 5-E).
Es ist deutlich zu erkennen, dass die geleistete Arbeit mit zunehmender innerer Kraft des Muskels zunimmt. Ab einer bestimmten Kraft steigt die geleistete Arbeit jedoch nicht weiter an, sondern bleibt konstant.
|
Beim Vergleich des Kraft-Geschwindigkeits-Diagramms (Abbildung
26) mit der Abbildung 4 fällt auf, daß
die Kontraktionsgeschwindigkeit nicht für verschwindende Kräfte
maximal wird wie bei der gestreiften Muskulatur, sondern nach
überschreiten einer maximalen Verkürzungsgeschwindigkeit auf Null
zurückgeht. Die maximale Verkürzungsgeschwindigkeit stellt sich
dann ein, wenn die aktuelle Muskellänge etwa das ![]() fache der
Ausgangslänge erreicht. Gleiches gilt für die maximalen
Verkürzungen gemäß Abbildung 25. Verkürzungen und
Verkürzungsgeschwindigkeiten verlaufen also quasi-affin in
Abhängigkeit von der Kraft.
fache der
Ausgangslänge erreicht. Gleiches gilt für die maximalen
Verkürzungen gemäß Abbildung 25. Verkürzungen und
Verkürzungsgeschwindigkeiten verlaufen also quasi-affin in
Abhängigkeit von der Kraft.
Hierraus kann geschlossen werden, dass die Kontraktilität -- gegeben durch die Anzahl der Querbrücken und ihrer ''Koppelfrequenz'' — von der Kraft und der Länge des Muskels abhängt. Trotz dieses grundsätzlich verschiedenen Verhaltens glatter und gestreifter Muskulatur, sind die Verläufe der geleisteten isotonischen Arbeiten ähnlich.